The Pristine Myth: The Landscape of the
Americas in 1492
William M. Denevan
Department of Geography, University of Wisconsin, Madison, WI 53706
Abstract. The myth persists that in 1492 the Americas were a sparsely populated wilderness, -a world of barely perceptible human disturbance.- There is substantial evidence, however, that the Native American landscape of the early sixteenth century was a humanized landscape almost everywhere. Populations were large. Forest composition had been modified, grasslands had been created, wildlife disrupted, and erosion was severe in places. Earthworks, roads, fields, and settlements were ubiquitous. With Indian depopulation in the wake of Old World disease, the environment recovered in many areas. A good argument can be made that the human presence was less visible in 1750 than it was in 1492.
Key Words: Pristine myth, 1492, Columbus, Native American settlement and demography, prehistoric New World, vegetation change, earthworks.
'This is the forest primeval ..."
Evangeline: A Tale of Acadie (Longfellow, 1847).
What was the New World like at the time of Columbus?-" Geography as it was," in the words of Carl Sauer (1971, x). The Admiral himself spoke of a 'Terrestrial Paradise," beautiful and green and fertile, teeming with birds, with naked people living there whom he called "Indians." But was the landscape encountered in the sixteenth century primarily pristine, virgin, a wilderness, nearly empty of people, or was it a humanized landscape, with the imprint of native Americans being dramatic and persistent? The former still seems to be the more common view, but the latter may be more accurate.
The pristine view is to a large extent an invention of nineteenth-century romanticist and primitivist writers such as W.H. Hudson, Cooper, Thoreau, Longfellow, and Parkman, and painters such as Catlin and Church! The wilderness image has since become part of the American heritage, associated *with a heroic pioneer past in need of preservation' (Pyne 1982, 17; also see Bowden 1992, 22). The pristine view was restated clearly in 1950 by John Bakeless in his book The Eyes of Discovery:
There were not really very many of these redmen ... the land seemed empty to invaders who came from settled Europe . . . that ancient, primeval, undisturbed wilderness . . . the streams simply boiled with fish ... so much game . . . that one hunter counted a thousand animals near a single salt lick ... the virgin wilderness of Kentucky ... the forested glory of primitive America (13, 201, 223, 314, 407).
But then he mentions that Indian "prairie fires . . . cause the often-mentioned oak openings ... Great fields of corn spread in all directions ... the Barrens ... without forest,' and that 'Early Ohio settlers found that they could drive about through the forests with sleds and horses" (31, 304, 308, 314). A contradiction?
In the ensuing forty years, scholarship has shown that Indian populations in the Americas were substantial, that the forests had indeed been altered, that landscape change was commonplace. This message, however, seems not to have reached the public through texts, essays, or talks by both academics and popularizers who have a responsibility to know better.3
Kirkpatrick Sale in 1990, in his widely reported Conquest of Paradise, maintains that it was the Europeans who transformed nature, following a pattern set by Columbus. Although Sale's book has some merit and he is aware of large Indian numbers and their impacts, he nonetheless champions the widely-held dichotomy of the benign Indian landscape and the devastated Colonial landscape. He overstates both.
Similarly, Seeds of Change: Christopher Columbus and the Columbian Legacy, the popular book published by the Smithsonian Institution, continues the litany of Native American passivity:
pre-Columbian America was still the First Eden, a pristine natural kingdom. The native people were transparent in the landscape, living as natural elements of the ecosphere. Their world, the New World of Columbus, was a world of barely perceptible human disturbance (Shetler 1991, 226).
To the contrary, the Indian impact was neither benign nor localized and ephemeral, nor were resources always used in a sound ecological way. The concern here is with the form and magnitude of environmental modification rather than with whether or not Indians lived in harmony with nature with sustainable systems of resource management. Sometimes they did; sometimes they didn't. What they did was to change their landscape nearly everywhere, not to the extent of post-Colonial Europeans but in important ways that merit attention.
The evidence is convincing. By 1492 Indian activity throughout the Americas had modified forest extent and composition, created and expanded grasslands, and rearranged microrelief via countless artificial earthworks. Agricultural fields were common, as were houses and towns and roads and trails. All of these had local impacts on soil, microclimate, hydrology, and wildlife. This is a large topic, for which this essay offers but an introduction to the issues, misconceptions, and residual problems. The evidence, pieced together from vague ethnohistorical accounts, field surveys, and archaeology, supports the hypothesis that the Indian landscape of 1492 had largely vanished by the mid-eighteenth century, not through a European superimposition, but because of the demise of the native population. The landscape of 1750 was more "pristine" (less humanized) than that of 1492.
Indian Numbers
The size of the native population at contact is critical to our argument. The prevailing position, a recent one, is that the Americas were well-populated rather than relatively empty lands in 1492. In the words of the sixteenth-century Spanish priest, Bartolomé de las Casas, who knew the Indies well:
All that has been discovered up to the year forty-nine 115491 is full of people, like a hive of bees, so that it seems as though God had placed all, or the greater part of the entire human race in these countries (Las Casas, in MacNutt 1909, 314).
Las Casas believed that more than 40 million Indians had died by the year 1560. Did he exaggerate? In the 1930s and 1940s, Alfred Kroeber, Angel Rosenblat, and Julian Steward believed that he had. The best counts then available indicated a population of between 815 million Indians in the Americas. Subsequently, Carl Sauer, Woodrow Borah, Sherburne F. Cook, Henry Dobyns, George Lovell, N. David Cook, myself, and others have argued for larger estimates. Many scholars now believe that there were between 40-100 million Indians in the hemisphere (Denevan 1992). This conclusion is primarily based on evidence of rapid early declines from epidemic disease prior to the first population counts (Lovell, this volume).
I have recently suggested a New World total of 53.9 million (Denevan 1992, xxvii). This divides into 3.8 million for North America, 17.2 million for Mexico, 5.6 million for Central America, 3.0 million for the Caribbean, 15.7 million for the Andes, and 8.6 million for lowland South America. These figures are based on my judgment as to the most reasonable recent tribal and regional estimates. Accepting a margin of error of about 20 percent, the New World population would lie between 43-65 million. Future regional revisions are likely to maintain the hemispheric total within this range. Other recent estimates, none based on totaling regional figures, include 43 million by Whitmore (1991, 483), 40 million by Lord and Burke (1991), 40-50 million by Cowley (1991), and 80 million for just Latin America by Schwerin (1991, 40). In any event, a population between 40-80 million is sufficient to dispel any notion of "empty lands." Moreover, the native impact on the landscape of 1492 reflected not only the population then but the cumulative effects of z growing population over the previous 15,000 years or more.
European entry into the New World abruptly reversed this trend. The decline of native American populations was rapid and severe, probably the greatest demographic disaster eve; (Lovell, this volume). Old World diseases were the primary killer. In many regions, particularly the tropical lowlands, populations fell by 90 percent or more in the first century after the contact. Indian populations (estimated) declined Hispaniola from 1 million in 1492 to a few hundred 50 years later, or by more than 99 percent in Peru from 9 million in 1520 to 670,000 in 1620 (92 percent); in the Basin of Mexico from 1.6 million in 1519 to 180,000 in 107 (89 percent); and in North America from 3.8 million in 1492 to 1 million in 1800 (74 percent). An overall drop from 53.9 million in 1492 to 5.6 million in 1650 amounts to an 89 percent reduction (Denevan 1992, xvii-xxix). The human landscape was affected accordingly, although there is not always a direct relationship between population density and human impact (Whitmore, et al. 1990, 37).
The replacement of Indians by Europeans and Africans was initially a slow process. By 1638 there were only about 30,000 English in North America (Sale 1990, 388), and by 1750 there were only 1.3 million Europeans and slaves (Meinig 1986, 247). For Latin America in 1750, 56nchez-Albornoz (1974, 7) gives a total (including Indians) of 12 million. For the hemisphere in 1750, the Atlas of World Population History reports 16 million (McEvedy and Jones 1978, 270). Thus the overall hemispheric population in 1750 was about 30 percent of what it may have been in 1492. The 1750 population, however, was very unevenly distributed, mainly located in certain coastal and highland areas with little Europeanization elsewhere. In North America in 1750, there were only small pockets of settlement beyond the coastal belt, stretching from New England to northern Florida (see maps in Meinig 1986, 209, 245). Elsewhere, combined Indian and European populations were sparse, and environmental impact was relatively minor.
Indigenous imprints on landscapes at the time of initial European contact varied regionally in form and intensity. Following are examples for vegetation and wildlife, agriculture, and the built landscape.
Vegetation
The Eastern Forests
The forests of New England, the Midwest, and the Southeast had been disturbed to varying degrees by Indian activity prior to European occupation. Agricultural clearing and burning had converted much of the forest into successional (fallow) growth and into semi-permanent grassy openings (meadows, barrens, plains, glades, savannas, prairies), often of considerable size.' Much of the mature forest was characterized by an open, herbaceous understory, reflecting frequent ground fires. "The de Soto expedition, consisting of many people, a large horse herd, and many swine, passed through ten states without difficulty of movement" (Sauer 1971, 283). The situation has been described in detail by Michael Williams in his recent history of American forests: "Much of the 'natural' forest remained, but the forest was not the vast, silent, unbroken, impenetrable and dense tangle of trees beloved by many writers in their romantic accounts of the forest wilderness" (1989, 33).' "The result was a forest of large, widely spaced trees, few shrubs, and much grass and herbage . . . Selective Indian burning thus promoted the mosaic quality of New England ecosystems, creating forests in many different states of ecological succession" (Cronon 1983, 49-51).
The extent, frequency, and impact of Indian burning is not without controversy. Raup (1937) argued that climatic change rather than Indian burning could account for certain vegetation changes. Emily Russell (1983, 86), assessing pre1700 information for the Northeast, concluded that: "There is no strong evidence that Indians purposely burned large areas," but Indians did "increase the frequency of fires above the low numbers caused by lightning," creating an open forest. But then Russell adds: "in most areas climate and soil probably played the major role in determining the precolonial forests." She regards Indian fires as mainly accidental and "merely" augmental to natural fires, and she discounts the reliability of many early accounts of burning.
Forman and Russell (1983, 5) expand the argument to North America in general: "regular and widespread Indian burning (Day 1953) [is] an unlikely hypothesis that regretfully has been accepted in the popular literature and consciousness." This conclusion, I believe, is unwarranted given reports of the extent of prehistoric human burning in North America and Australia (Lewis 1982), and Europe (Patterson and Sassaman 1988, 130), and by my own and other observations on current Indian and peasant burning in Central America and South America; when unrestrained, people burn frequently and for many reasons. For the Northeast, Patterson and Sassaman (1988 ' 129) found that sedimentary charcoal accumulations were greatest where Indian populations were greatest.
Elsewhere in North America, the Southeast is much more fire prone than is the Northeast, with human ignitions being especially important in winter (Taylor 1981). The Berkeley geographer and Indianist Erhard Rostlund (1957, 1960) argued that Indian clearing and burning created many grasslands within mostly open forest in the so-called "prairie belt" of Alabama. As improbable as it may seem, Lewis (1982) found Indian burning in the subarctic, and Dobyns (1981) in the Sonoran desert. The characteristics and impacts of fires set by Indians varied regionally and locally with demography, resource management techniques, and environment, but such fires clearly had different vegetation impacts than did natural fires owing to differences in frequency, regularity, and seasonality.
Forest Composition
In North America, burning not only maintained open forest and small meadows but also encouraged fire-tolerant and sun-loving species. 'Fire created conditions favorable to strawberries, blackberries, raspberries, and other gatherable foods" (Cronon 1983, 51). Other useful plants were saved, protected, planted, and transplanted, such as American chestnut, Canada plum, Kentucky coffee tree, groundnut, and leek (Day 1953, 339-40). Gilmore (1931) described the dispersal of several native plants by Indians. Mixed stands were converted to single species dominants, including various pines and oaks, sequoia, Douglas fir, spruce, and aspen (M. Williams 1989, 4748). The longleaf, slash pine, and scrub oak forests of the Southeast are almost certainly an anthropogenic subclimax created originally by Indian burning, replaced in early Colonial times by mixed hardwoods, and maintained in part by fires set by subsequent farmers and woodlot owners (Garren 1943). Lightning fires can account for some fire-climax vegetation, but Indian burning would have extended and maintained such vegetation (Silver 1990, 17-19, 59--64).
Even in the humid tropics, where natural fires are rare, human fires can dramatically influence forest composition. A good example is the pine forests of Nicaragua (Denevan 1961). Open pine stands occur both in the northern highlands (below 5,000 feet) and in the eastern (Miskito) lowlands, where warm temperatures and heavy rainfall generally favor mixed tropical montane forest or rainforest. The extensive pine forests of Guatemala and Mexico primarily grow in cooler and drier, higher elevations, where they are in large part natural and prehuman (Watts and Bradbury 1982, 59). Pine forests were definitely present in Nicaragua when Europeans arrived. They were found in areas where Indian settlement was substantial, but not in the eastern mountains where Indian densities were sparse. The eastern boundary of the highland pines seems to have moved with an eastern settlement frontier that has fluctuated back and forth since prehistory. The pines occur today where there has been clearing followed by regular burning and the same is likely in the past. The Nicaraguan pines are fire tolerant once mature, and large numbers of seedlings survive to maturity if they can escape fire during their first three to seven years (Denevan 1961, 280). Where settlement has been abandoned and fire ceases, mixed hardwoods gradually replace pines. This succession is likely similar where pines occur elsewhere at low elevations in tropical Central America, the Caribbean, and Mexico.
Midwest Prairies and Tropical Savannas
Sauer (1950, 1958, 1975) argued early and often that the great grasslands and savannas of the New World were of anthropogenic rather than climatic origin, that rainfall was generally sufficient to support trees. Even nonagricultural Indians expanded what may have been pockets of natural, edaphic grasslands at the expense of forest. A fire burning to the edge of a grass/forest boundary will penetrate the drier forest, margin and push back the edge, even if the forest itself is not consumed (MuellerDombois 1981, 164). Grassland can therefore advance significantly in the wake of hundreds of years of annual fires. Lightning-set fires can have a similar impact, but more slowly if less frequent than human fires, as in the wet tropics.
The thesis of prairies as fire induced, primarily by Indians, has its critics (Borchert 1950; Wedel 1957), but the recent review of the topic by Anderson (1990, 14), a biologist, concludes that most ecologists now believe that the eastern prairies "would have mostly disappeared if it had not been for the nearly annual burning of these grasslands by the North American Indians," during the last 5,000 years. A case in point is the nineteenth-century invasion of many grasslands by forests after fire had been suppressed in Wisconsin, Illinois, Kansas, Nebraska, and elsewhere (M. Williams 1989,46).
The large savannas of South America are also controversial as to origin. Much, if not most of the open vegetation of the Orinoco Llanos, the Llanos de Mojos of Bolivia, the Pantanal of Moto Grosso, the Bolfvar savannas of Colombo, the Guayas savannas of coastal Ecuador, the campo cerrado of central Brazil, and the coastal savannas north of the Amazon, is of natural origin. The vast campos; cerrados; occupy extremely senile, often toxic oxisols. The seasonally inundated savannas of Bolivia, Brazil, Guayas, and the Orinoco owe their existence to the intolerance of woody species to the extreme alternation of lengthy flooding or waterlogging and severe desiccation during a long dry season. These savannas, however, were and are burned by Indians and ranchers, and such fires have expanded the savannas into the forests to an unknown extent. It is now very difficult to determine where a natural forest/savanna boundary once was located (Hills and Randall 1968; Medina 1980).
Other small savannas have been cut out of the rainforest by Indian farmers and then maintained by burning. An example is the Gran Pajonal in the Andean foothills in east-central Peru, where dozens of small grasslands (pajonales) have been created by Campa Indians-a process clearly documented by air photos (Scott 1978). Pajonales were in existence when the region was first penetrated by Franciscan missionary explorers in 1733.
The impact of human activity is nicely illustrated by vegetational changes in the basins of the San Jorge, Cauca, and Sinú rivers of northern Colombia. The southern sector, which was mainly savanna when first observed in the sixteenth century, had reverted to rainforest by about 1750 following Indian decline, and had been reconverted to savanna for pasture by 1950 (Gordon 1957, map p. 69). Sauer (1%6, 285-M; 1976, 8) and Bennett (1968, 53-55) cite early descriptions of numerous savannas in Panama in the sixteenth century. Balboa's first view of the Pacific was from a "treeless ridge," now probably forested. Indian settlement and agricultural fields were common at the time, and with their decline the rainforest returned.
Anthropogenic Tropical Rain Forest
The tropical rain forest has long had a reputation for being pristine, whether in 1492 or 1992. There is, however, increasing evidence that the forests of Amazonia and elsewhere are largely anthropogenic in form and composition. Sauer (1958, 105) said as much at the Ninth Pacific Science Congress in 1957 when he challenged the statement of tropical botanist Paul Richards that, until recently, the tropical forests have been largely uninhabited, and that prehistoric people had 'no more influence on the vegetation than any of the other animal inhabitants." Sauer countered that Indian burning, swiddens, and manipulation of composition had extensively modified the tropical forest.
"Indeed, in much of Amazonia, it is difficult to find soils that are not studded with charcoal" (Uhl, et al. 1990, 30). The question is, to what extent does this evidence reflect Indian burning in contrast to natural (lightning) fires, and when did these fires occur? The role of fire in tropical forest ecosystems has received considerable attention in recent years, partly as result of major wild fires in East Kalimantan in 1982-83 and small forest fires in the Venezuelan Amazon in 1980-84 (Goidammer 1990). Lightning fires, though rare in moist tropical forest, do occur in drier tropical woodlands (MuellerDombois 1981, 149). Thunderstorms with lightning are much more common in the Amazon, compared to North America, but in the tropics lightning is usually associated with heavy rain and noncombustible, verdant vegetation. Hence Indian fires undoubtedly account for most fires in prehistory, with their impact varying with the degree of aridity.
In the Rio Negro region of the Colombian-Venezuelan Amazon, soil charcoal is very common in upland forests. C-14 dates range from 6260-250 B.P., well within human times (Saldarriaga and West 1986). Most of the charcoal probably reflects local swidden burns; however, there are some indications of forest fires at intervals of several hundred years, most likely ignited by swidden fires. Recent wild fires in the upper Rfo Negro region were in a normally moist tropical forest (3530 mrn annual rainfall) that had experienced several years of severe drought. Such infrequent wild fires in prehistory, along with the more frequent ground fires, could have had significant impacts on forest succession, structure, and composition. Examples are the pine forests of Nicaragua, mentioned above, the oak forests of Central America, and the babassu palm forests of eastern Brazil. Widespread and frequent burning may have brought about the extinction of some endemic species.
The Amazon forest is a mosaic of different ages, structure, and composition resulting from local habitat conditions and disturbance dynamics (Haffer 1991). Natural disturbances (tree falls, landslides, river activity) have been considerably augmented by human activity, particularly by shifting cultivation. Even a small number of swidden farmers can have a widespread impact in a relatively short period of time. In the Rio Negro region, species-diversity recovery takes 60-W years and biomass recovery 140-200 years (Saldarriaga and Uhl 1991, 312). Brown and Lugo (1990, 4) estimate that today about forty percent of the tropical forest in Latin America is secondary as a result of human clearing and that most of the remainder has had some modification despite current low population densities. The species composition of early stages of swidden fallows differs from that of natural gaps and may "alter the species composition of the mature forest on a longterm scale" (WaIschburger and Von Hildebrand 1991, 262). While human environmental destruction in Amazonia currently is concentrated along roads, in prehistoric times Indian activity in the upland (interflueve) forests was much less intense but more widespread (Denevan forthcoming).
Indian modification of tropical forests is not limited to clearing and burning. Large expanses of Latin American forests are humanized forests in which the kinds, numbers, and distributions of useful species are managed by human populations. Doubtless, this applies to the past as well. One important mechanism in forest management is manipulation of swidden fallows (sequential agroforestry) to increase useful species. The planting, transplanting, sparing, and protection of useful wild, fallow plants eliminates clear distinctions between field and fallow (Denevan and Padoch 1988). Abandonment is a slow process, not an event. Gordon (1982, 79-98) describes managed regrowth vegetation in eastern Panama, which he believes extended from Yucat6n to northern Colombia in pre-European times. The Huastec of eastern Mexico and the Yucatec Maya have similar forms of forest gardens or forest management (Alcorn 1981; G6mez-Pompa 1987). The Kayap6 of the Brazilian Amazon introduce and/or protect useful plants in activity areas ("nomadic agriculture") adjacent to villages or camp sites, in foraging areas, along trails, near fields, and in artificial forest-mounds in savanna (Posey 1985). In managed forests, both annuals and perennials are planted or transplanted, while wild fruit trees are particularly common in early successional growth. Weeding by hand was potentially more selective than indiscriminate weeding by machete (Gordon 1982, 57-61). Much dispersal of edible plant seeds is unintentional via defecation and spitting out.
The economic botanist William Bal6e (1987, 1989) speaks of "cultural" or "anthropogenic" forests in Amazonia in which species have been manipulated, often without a reduction in natural diversity. These include specialized forests (babassu, Brazil nuts, lianas, palms, bamboo), which currently make up at least 11.8 percent (measured) of the total upland forest in the Brazilian Amazon (Bal6e 1989,14). Clear indications of past disturbance are the extensive zones of terra preta (black earth), which occur along the edges of the large floodplains as well as in the uplands (Bal6e 1989, 10-12; Smith 1980). These soils, with depths to 50 cm or more, contain charcoal and cultural waste from prehistoric burning and settlement. Given high carbon, nitrogen, calcium, and phosphorus content, terra preta soils have a distinctive vegetation and are attractive to farmers. Bal~e (1989, 14) concludes that "large portions of Amazonian forests appear to exhibit the continuing effects of past human interference." The same argument has been made for the Kenya lowlands (Gómez--Pompa, et al. 1987) and Panama (Gordon, 1982). There are no virgin tropical forests today, nor were there in 1492.
Wildlife
The indigenous impact on wildlife is equivocal. The thesis that "overkill" hunting caused the extinction of some large mammals in North America during the late Pleistocene, as well as subsequent local and regional depletions (Martin 1978,167-72), remains controversial. By the time of the arrival of Cortéz in 1519, the dense populations of Central Mexico apparently had greatly reduced the number of large game, given reports that "they eat any living thing" (Cook and Borah 1971-79, (3) 135, 140). In Amazonia, local game depletion apparently increases with village size and duration (Good 1987). Hunting procedures in many regions seem, however, to have allowed for recovery because of the 'resting" of hunting zones intentionally or as a result of shifting of village sites.
On the other hand, forest disturbance increased herbaceous forage and edge effect, and hence the numbers of some animals (Thompson and Smith 1970, 261-4A). "Indians created ideal habitats for a host of wildlife species . . . exactly those species whose abundance so impressed English colonists: elk, deer, beaver, hare, porcupine, turkey, quail, ruffed grouse, and so on" (Cronon 1983, 51). Mite-tailed deer, peccary, birds, and other game increases in swiddens and fallows in Yucatán and Panama (Greenberg 1991; Gordon 1982, 96-112; Bennett 1968). Rostlund (1960, 407) believed that the creation of grassy openings east of the Mississippi extended the range of the bison, whose numbers increased with Indian depopulation and reduced hunting pressure between 1540-1700, and subsequently declined under White pressure.
Agriculture
Fields and Associated Features
To observers in the sixteenth century, the most visible manifestation of the Native American landscape must have been the cultivated fields, which were concentrated around villages and houses. Most fields are ephemeral, their presence quickly erased when farmers migrate or die, but there are many eye-witness accounts of the great extent of Indian fields. On Hispaniola, Las Casas and Oviedo reported individual fields with thousands of montones (Sturtevant 1961, 73j. These were manioc and sweet potato mounds 3-4 m in circumference, of which apparently none have survived. In the Llanos de Moios in Bolivia, the first explorers mentioned percheles, or corn cribs on pilings, numbering up to 700 in a single field, each holding 30-45 bushels of food (Denevan 1966, 98). In northern Florida in 1539, Hernando de Soto's army passed through numerous fields of maize, beans, and squash, their main source of provisions; in one sector, 'great fields . . . were spread out as far as the eye could see across two leagues of the plainff (Garcilaso de la Vega 1980, (2) 182; also see Dobyns 1983, 135-46).
It is difficult to obtain a reliable overview from such descriptions. Aside from possible exaggeration, Europeans tended not to write about field size, production, or technology. More useful are various forms of relict fields and field features that persist for centuries and can still be recognized, measured, and excavated today. These extant features, including terraces, irrigation works, raised fields, sunken fields, drainage ditches, dams, reservoirs, diversion walls, and field borders number in the millions and are distributed throughout the Americas (Denevan 1980; see also Doolittle and Whitmore and Turner, this volume). For example, about 500,000 ha of abandoned raised fields survive in the San Jorge Basin of northern Colombia (Plazas and Falchetti 1987, 485), and at least 600,000 ha of terracing, mostly of prehistoric origin, occur in the Peruvian Andes (Denevan 1988, 20). There are 19,000 ha of visible raised fields in just the sustaining area of Tiwanaku at Lake Titicaca (Kolata 1991,109) and there were about 12,000 ha of chinampas (raised fields) around the Aztec capital of TenochtitlAn (Sanders, et al. 1979, 390). Complex canal systems on the north coast of Peru and in the Salt River Valley in Arizona irrigated more land in pre~istory than is cultivated today. About 175 sites of Indian garden beds, up to several hundred acres each, have been reported in Wisconsin (Gartner 1992). These various remnant fields probably represent less than 25 percent of what once existed, most being buried under sediment or destroyed by erosion, urbanization, plowing, and bulldozing. On the other hand, an inadequate effort has been made to search for ancient fields.
Erosion
The size of native populations, associated deforestation, and prolonged intensive agriculture led to severe land degradation in some regions. Such a landscape was that of Central Mexico, where by 1519 food production pressures may have brought the Aztec civilization to the verge of collapse even without Spanish intervention (Cook and Borah 1971-79 (3), 12976).' There is good evidence that severe soil erosion was already widespread, rather than just the result of subsequent European plowing, livestock, and deforestation. Cook examined the association between erosional severity (gullies, barrancas, sand and silt deposits, and sheet erosion) and pre-Spanish population density or proximity to prehistoric Indian towns. He concluded that "an important cycle of erosion and deposition therefore accompanied intensive land use by huge primitive populations in central Mexico, and had gone far toward the devastation of the country before the white man arrived" (Cook 1949, 86).
Barbara Williams (1972, 618) describes widespread tepetate, an indurated substrate formation exposed by sheet erosion resulting from prehistoric agriculture, as 0one of the dominant surface materials in the Valley of Mexico.' On the other hand, anthropologist Melville (1990, 27) argues that soil erosion in the Valle de Mezquital, just north of the Valley of Mexico, was the result of overgrazing by Spanish livestock starting before 1600: "there is an almost total lack of evidence of environmental degradation before the last three decades of the sixteenth century." The Butzers, however, in an examination of Spanish land grants, grazing patterns, and soil and vegetation ecology, found that there was only light intrusion of Spanish livestock (sheep and cattle were moved frequently) into the southeastern Baifo near Mezquital until after 1590 and that any degradation in 1590 was "as much a matter of long-term Indian land use as it was of Spanish intrusion' (Butzer and Butzer forthcoming). The relative roles of Indian and early Spanish impacts in Mexico still need resolution; both were clearly significant but varied in time and place. Under the Spaniards, however, even with a greatly reduced population, the landscape in Mexico generally did not recover due to accelerating impacts from introduced sheep and cattle!
The Built Landscape
Settlement
The Spaniards and other Europeans were impressed by large flourishing Indian cities such as Tenochtitlán, Quito, and Cuzco, and they took note of the extensive ruins of older, abandoned cities such as Cahokia, TeotihuacAn, T ikal, Chan Chan, and Tiwanaku (Harcloy 1968). Most of these cities contained more than 50,000 people. Less notable, or possibly more taken for granted, was rural settlement-small villages of a few thousand or a few hundred people, hamlets of a few families, and dispersed farmsteads. The numbers and locations of much of this settlement will never be known. With the rapid decline of native populations, the abandonment of houses and entire villages and the decay of perishable materials quickly obscured sites, especially in the tropical lowlands.
We do have some early listings of villages, especially for Mexico and Peru. Elsewhere, archaeology is telling us more than ethnohistory. After initially focusing on large temple and administrative centers, archaeologists are now examining rural sustaining areas, with remarkable results. See, for example, Sanders et al. (1979) on the Basin of Mexico, Culbert and Rice (1991) on the Maya lowlands, and Fowler (1989) on Cahokia in Illinois. Evidence of human occupation for the artistic Santar6m Culture phase (Tapaj6s chiefdom) on the lower Amazon extends over thousands of square kilometers, with large nucleated settlements (Roosevelt 1991, 101-02).
Much of the rural precontact settlement was semi-dispersed (rancherias), particularly in densely populated regions of Mexico and the Andes, probably reflecting poor food transport efficiency. Houses were both single-family and communal (pueblos, Huron long houses, Amazon malocas). Construction was of stone, earth, adobe, daub and wattle, grass, hides, brush, and bark. much of the dispersed settlement not destroyed by depopulation was concentrated by the Spaniards into compact and plaza style new towns (congregaciones, reducciones) for administrative purposes.
Mounds
James Parsons (1985, 161) has suggested that: "-An apparent 'mania for earth moving, landscape engineering on a grand scale runs as a thread through much of New World prehistory. Large quantities of both earth and stone were transferred to create various raised and sunken features, such as agricultural landforms, settlement and ritual mounds, and causeways.
Mounds of different shapes and sizes were constructed throughout the Americas for temples, burials, settlement, and as effigies. The stone pyramids of Mexico and the Andes are well known, but equal monuments of earth were built in the Amazon, the Midwest U.S., and elsewhere. The Mississippian period complex of 104 mounds at Cahokia near East St. Louis supported 30,000 people; the largest, monk's Mound, is currently 30.5 m high and covers 6.9 ha. (Fowler 1989, 90, 192). Cahokia was the largest settlement north of the Rio Grande until surpassed by New York City in 1775. An early survey estimated "at least 20,000 conical, linear, and effigy mounds" in Wisconsin (Stout 1911, 24). Overall, there must have been several hundred thousand artificial mounds in the Midwest and South. De Soto described such features still in use in 1539 (Silverberg 1968, 7). Thousands of settlement and other mounds dot the savanna landscape of Mojos in Bolivia (Denevan 1966). At the mouth of the Amazon on Marajó Island, one complex of forty habitation mounds contained more than 10,000 people; one of these mounds is 20 m high while another is 90 ha in area (Roosevelt 1991, 31, 38).
Not all of the various earthworks scattered over the Americas were in use in 1492. Many had been long abandoned, but they constituted a conspicuous element of the landscape of 1492 and some are still prominent. Doubtless, many remain to be discovered, and others remain unrecognized as human or prehistoric features.
Roads, Causeways, and Trails
Large numbers of people and settlements necessitated extensive systems of overland travel routes to facilitate administration, trade, warfare, and social interaction (Hyslop 1984; Trombold 1991). Only hints of their former prominence survive. Many were simple traces across deserts or narrow paths cut into forests. A suggestion as to the importance of Amazon forest trails is the existence of more than 500 krin of trail maintained by a single Kayap6 village today (Posey 1985, 149). Some prehistoric footpaths were so intensively used for so long that they were incised into the ground and are still detectable, as has recently been described in Costa Rica (Sheets and Sever 1991).
Improved roads, at times stone-lined and drained, were constructed over great distances in the realms of the high civilizations. The Inca road network is estimated to have measured about 40,000 km, extending from southern Colombia to central Chile (Hyslop 1984, 224). Prehistoric causeways (raised roads) were built in the tropical lowlands (Denevan 1991); one Maya causeway is 100 krn long, and there are more than 1,600 km of causeways in the Lianos de Mojos. Humboldt reported large prehistoric causeways in the Orinoco Llanos. Ferdinand Columbus described roads on Puerto Rico in 1493. Gaspar de Carvajal, traveling down the Amazon with Orellana in 1541, reported "highways" penetrating the forest from river bank villages. Joseph de Acosta (1880, (1) 171) in 1590 said that between Peru and Brazil, there were "waies as much beaten as those betwixt Salamanca and Valladolid." Prehistoric roads in Chaco Canyon, New Mexico are described in Trombold (1991). Some routes were so well established and located that they have remained roads to this day.
Recovery
A strong case can be made for significant environmental recovery and reduction of cultural features by the late eighteenth century as a result of Indian population decline. Henry Thoreau (1949, 132-37) believed, based on his reading of William Wood, that the New England forests of 1633 were more open, more park-like, with more berries and more wildlife, than Thoreau observed in 1855. Cronon (1983, 108), Pyne (1982, 51), Silver (1990, 104), Martin (1978,181-82), and Williams (1989, 49) all maintain that the eastern forests recovered and filled in as a result of Indian depopulation, field abandonment, and reduction in burning. While probably correct, these writers give few specific examples, so further research is needed. The sixteenth-century fields and sa' vannas of Colombia and Central America also had reverted to forest within 150 years after abandonment (Parsons 1975, 30-31; Bennett 1968, 54). On his fourth voyage in 1502-03, Columbus sailed along the north coast of Panama (Veragua). His son Ferdinand described lands which were well-peopled, full of houses, with many fields, and open with few trees. In contrast, in 1681 Lionel Wafer found most of the Caribbean coast of Panama forest covered and unpopulated. On the Pacific side in the eighteenth century, savannas were seldom mentioned; the main economic activity was the logging of tropical cedar, a tree that grows on the sites of abandoned fields and other disturbances (Sauer 1966, 132-33, 287-M). An earlier oscillation from forest destruction to recovery in the Yucatán is instructive. Whitmore, et al. (1990, 35) estimate that the Maya had modified 75 percent of the environment by A.D. 800, and that following the Mayan collapse, forest recovery in the central lowlands was nearly complete when the Spaniards arrived.
The pace of forest regeneration, however, varied across the New World. Much of the southeastern U.S. remained treeless in the 1750s according to Rostlund (1957, 408, 409). He notes that the tangled brush that ensnarled the "Wilderness Campaign of 1864 in Virginia occupied the same land as did Captain John Smith's 'open groves with much good ground between without any shrubs" in 1624; vegetation had only partially recovered over 240 years. The Kentucky barrens in contrast were largely reforested by the early nineteenth century (Sauer 1963, 30). The Alabama Black Belt vegetation was described by William Bartrarn in the 1770s as a mixture of forest and grassy plains, but by the nineteenth century, there was only 10 percent prairie and even less in some counties (Rostlund 1957, 393, 401-03). Sections of coastal forests never recovered, given colonist pressures, but Sale's (1990, 291) claim that 'the English were well along in the process of eliminating the ancient Eastern woodlands from Maine to the Mississippi" in the first one hundred years, is an exaggeration.
Wildlife also partially recovered in eastern North America with reduced hunting pressure from Indians; however, this is also a story yet to be worked out. The white-tailed deer apparently declined in numbers, probably reflecting reforestation plus competition from livestock. Commercial hunting was a factor on the coast, with 80,000 deer skins being shipped out yearly from Charleston by 1730 (Silver 1990, 92). Massachusetts enacted a closed season on deer as early as 1694, and in 1718 there was a three-year moratorium on deer hunting (Cronon 1983, 100). Sale (1990, 290) believes that beaver were depleted in the Northeast by 1640. Other fur bearers, game birds, elk, buffalo, and carnivores were also targeted by white hunters, but much game probably was in the process of recovery in many eastern areas until a general reversal after 1700-50.
As agricultural fields changed to scrub and forest, earthworks were grown over. All the raised fields in Yucatán and South America were abandoned. A large portion of the agricultural terraces in the Americas were abandoned in the early colonial period (Donkin 1979, 35-38). In the Colca Valley of Peru, measurement on air photos indicates 61 percent terrace abandonment (Denevan 1988, 28). Societies vanished or declined everywhere and whole villages with them. The degree to which settlement features were swallowed up by vegetation, sediment, and erosion is indicated by the difficulty of finding them today. Machu Picchu, a late prehistoric site, was not rediscovered until 1911.
The renewal of human impact also varied regionally, coming with the Revolutionary War in North America, with the rubber boom in Amazonia, and with the expansion of coffee in southern Brazil (1840-1930). The swamp lands of Gulf Coast Mexico and the Guayas Basin of Ecuador remained hostile environments to Europeans until well into the nineteenth century or later (Siemens 1990; Mathewson 1987). On the other hand, Highland Mexico-Guatemala and the Andes, with greater Indian survival and with the establishment of haciendas and intensive mining, show less evidence of environmental recovery. Similarly, Indian fields in the Caribbean were rapidly replaced by European livestock and sugar plantation systems, inhibiting of recent recovery. The same is true of the sugar zone of coastal Brazil.
Conclusions
In 1492, Indian activity had modified vegetation and wildlife, caused erosion, and created earthworks, roads, and settlements throughout the Americas. This may be obvious, but the human imprint was much more ubiquitous and enduring than is usually realized. The historical evidence is ample, as are data from surviving earthworks and archaeology. And much can be inferred from present human impacts. The weight of evidence suggests that Indian populations were large, not only in Mexico and the Andes, but also in seemingly unattractive habitats such as the rainforests of Amazonia, the swamps of Mojos, and the deserts of Arizona.
Clearly, the most humanized landscapes of the Americas existed in those highland regions where people were the most numerous. Here were the large states, characterized by urban (enters, road systems, intensive agriculture, a dispersed but relatively dense rural settlement pattern of hamlets and farmsteads, and widespread vegetation and soil modification and wildlife depletion. There were other, smaller regions that shared some of these characteristics, such as the Pueblo lands in the southwestern U.S., the Sabana cle Bogotá in highland Colombia, and the central Amazon floodplain, where built landscapes were locally dramatic and are still observable. Finally, there were the immense grasslands, deserts, mountains, and forests elsewhere, with populations that were sparse or moderate, with landscape impacts that mostly were ephemeral or not obvious but nevertheless significant, particularly for vegetation and wildlife, as in Amazonia and the northeastern U.S. In addition, landscapes from the more distant past survived to 1492 and even to 1992, such as those of the irrigation states of north coast Peru, the Classic Maya, the Mississippian mound builders, and the Tiwanaku Empire of Lake Titicaca.
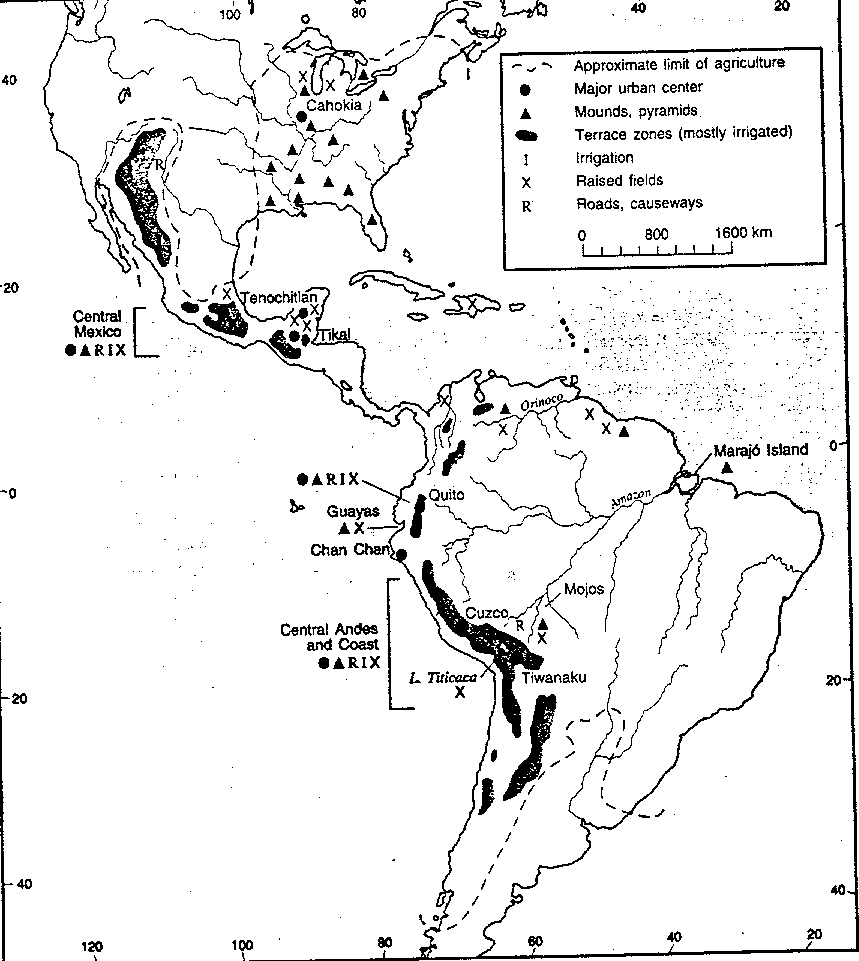
This essay has ranged over the hemisphere, an enormous area, making generalizations about and providing examples of Indian landscape transformation as of 1492. Examples of some of the surviving cultural features are shown in Figure 1. Ideally, a series of hemispheric maps should be provided to portray the spatial patterns of the different types of impacts and cultural features, but such maps are not feasible nor would they be accurate given present knowledge. There are a few relevant regional maps, however, that can be referred to. For example, see Butzer (1990, 33, 45) for Indian settlement structures/mounds and subsistence patterns in the U.S.; Donkin (1979, 23) for agricultural terracing; Doolittle (1990, 109) for canal irrigation in Mexico; Parsons and Denevan (1967) for raised fields in South America; Trombold (1991) for various road networks; Hyslop (1984, 4) for the Inca roads; Harcloy (1968, 49) for the most intense urbanization in Latin America; and Gordon (1957, 69) for anthropogenic savannas in northern Colombia.
The pristine myth cannot be laid at the feet of Columbus. While he spoke of 'Paradise," his was clearly a humanized paradise. He described Hispaniola and Tortuga as densely populated and "completely cultivated like the countryside around Cordoba" (Colón 1976, 165). He also noted that "the islands are not so thickly wooded as to be impassable,' suggesting openings from clearing and burn ing (Columbus 1961, 5).
The roots of the pristine myth lie in part with early observers unaware of human impacts that may be obvious to scholars today, particularly for vegetation and wildlife! But even many earthworks such as raised fields have only recently been discovered (Denevan 1966; 1980). Equally important, most of our eyewitness descriptions of wilderness and empty lands come from a later time, particularly 1750-1850 when interior lands began to be explored and occupied by Europeans. By 1650, Indian populations in the hemisphere had been reduced by about 90 percent, while by 1750 European numbers were not yet substantial and settlement had only begun to expand. As a result, fields had been abandoned. while settlements vanished, forests recovered. and savannas retreated. The landscape did appear to be a sparsely populated wilderness. This is the image conveyed by Parkman in the nineteenth century, Bakeless in 1950, and Shetler as recently as 1991. There was some-European impact, of course, but it was localized. After 1-50 and especially after 1850, populations greatly expanded, resources were more intensively exploited, and European modification of the environment accelerated, continuing to the present.
Figure 1. Selected features of the prehistoric cultural landscape. Some cities and agricultural works had been abandoned by 1492. The approximate limit of agriculture and the distribution of terraces is based on Donkin (1979, 23); other features were mapped by the author.
It is possible to conclude not only that "the virgin forest was not encountered in the sixteenth and seventeenth centuries; [but that] it was invented in the late eighteenth and early nineteenth centuries" (Pyne 1982, 46). However, "paradoxical as it may seem, there was undoubtedly much more 'forest primeval' in1850 than in 1650" (Rostlund 1957, 409). Thus the "invention" of an earlier wilderness is in part understandable and is not simply a deliberate creation which ennobled the American enterprise, as suggested by Bowden (1992 ' 2023). In any event, while pre-European landscape alteration has been demonstrated previously including by several geographers, the case has mainly been made for vegetation and mainly for eastern North America. As shown here, the argument is also applicable to most of the rest of the New World, including the humid tropics, and involves much more than vegetation.
The human impact on environment is not simply a process of increasing change or degradation in response to linear population growth and economic expansion. It is instead interrupted by periods of reversal and ecological rehabilitation as cultures collapse, populations decline, wars occur, and habitats are abandoned. Impacts may be constructive, benign, or degenerative (all subjective concepts), but change is continual at variable rates and in different directions. Even mild impact s and slow changes are cumulative, and the long-term effects can be dramatic. Is it possible that the thousands of years of human activity before Columbus created more change in the visible landscape than has occurred subsequently with European settlement and resource exploitation? The answer is probably yes for most regions for the next 250 years or so, and for some regions right up to the present time. American flora, fauna, and landscape were slowly Europeanized after 1492, but before that they had already been Indianized. "It is upon this imprint that the more familiar Euro-American landscape was grafted, rather than created anew" (Butzer 1990, 28). What does all this mean for protectionist tendencies today? Much of what is protected or proposed to be protected from human disturbance had native people present, and environmental modification occurred accordingly and in part is still detectable.
The pristine image of 1492 seems to be a myth, then, an image more applicable to 1750, following Indian decline, although recovery had only been partial by that date. There is some substance to this argument, and it should hold up under the scrutiny of further investigation of the considerable evidence available, both written and in the ground.
Acknowledgments
The field and library research that provided the background for this essay was undertaken over many years in Latin America, Berkeley, and Madison. Mentors who have been particularly influential are Carl 0. Sauer, Erhard Rostlund, James J. Parsons, and Woodrow Borah, all investigators of topics discussed
here.
Notes
References