 In addition to using amino acid racemization for geochronology, a primary goal of the Amino Acid Geochronology Laboratory is to use racemization to estimate the magnitude of late Quaternary paleotemperature changes. Unlike many other proxy climate recorders, temperature-dependent chemical reactions that take place in fossils are independent of paleoprecipitation changes, and therefore allow paleotemperatures to be calculated uniquely.
In addition to using amino acid racemization for geochronology, a primary goal of the Amino Acid Geochronology Laboratory is to use racemization to estimate the magnitude of late Quaternary paleotemperature changes. Unlike many other proxy climate recorders, temperature-dependent chemical reactions that take place in fossils are independent of paleoprecipitation changes, and therefore allow paleotemperatures to be calculated uniquely.
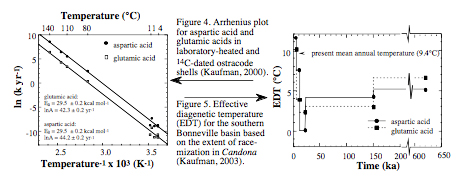
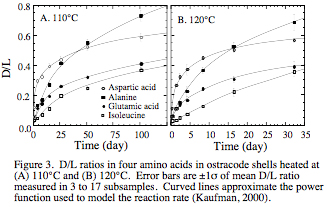
First, I developed equations that described the temperature sensitivity of the rate of amino acid racemization (Kaufman, 2000). I performed an extensive suite of heating experiments to evaluate the rate of the reaction over a range of temperatures (80 to 140°C) and D/L ratios (~0.1 to 0.6 in aspartic acid) in the common ostracode genus, Candona (Fig. 3). For each series of D/L ratios under isothermal conditions, I modeled the forward rate constant for the racemization using an empirical power-law model that approaches an asymptote of1.0. In addition to data from the heating experiments, the evaluation of racemization kinetics was extended to ambient temperatures using unheated, fossil ostracode shells from 14C-dated Holocene samples. Rate constants for both the heated and fossil samples were combined to calculate the Arrhenius parameters (activation energy and frequency factor) for racemization of two amino acids (aspartic acid and glutamic acid) (Fig. 4). This resulted in an uncalibrated age equation that relates sample D/L to the average postdepositional temperature and the sample age.