INTRODUCTION TO IMMUNITY
The UNDERLYING UNIVERSAL PRINCIPLE of the immune
system deals with recognizing (FOREIGN)
based on the principle of ligand/receptor binding described the
figure above. In a competitive and deadly world, self is usually GOOD
and nonself is usually BAD.
How is SELF recognized? How is NONSELF
(foreign) recognized?
We CAN NOT SURVIVE without a functioning immune
system. Without it, no amount of antibiotics or medical treatment can keep
us alive for more than a brief time. This is painfully illustrated by the
death of AIDS victims.
THE SPECIFIC IMMUNE SYSTEM = Previously
we have discussed the nonspecific defense system that protects us, more
or less, from all pathogens. The specific immune system (or often called
the IMMUNE SYSTEM) protects us against SPECIFIC NONSELF ORGANISMS and substances.
It is an INDUCED response; that is it must be TAUGHT which things to attack.
ANTIGEN = An antigen is anything that elicits
the
formation of a specific immune response. Older definitions limits the definition
of an antigen to ".....formation of an antibody.", however,
as you will learn there are two levels (duality) to the immune system.
EPITOPES = These are the unique regions
(or chemical groups) on a molecule that are antigenic; i.e., that elicit
a specific immune response.
ANTIBODY = A special group of soluble
proteins that are produced in response to foreign antigens.
IMMUNE CELLS or LYPHOCYTES= These
are the VARIOUS CELLS of the specific immunity system that respond
to SPECIFIC foreign or nonself antigens.
Antibodies are a group of soluble, proteins
that
have unique binding sites on them which recognize and bind to the
epitopes
of antigens. As previously described with enzymes, allosteric sites and
other binding site-situations, the antibody binding sites are
higly
specific. There are several types of antibodies with a variety
of different functions in the specific immune response which will be discussed
as appropriate. Figure 1 illustrates the relationship between an antigenic
molecule, its epitopes and the soluble antibodies produced against it.
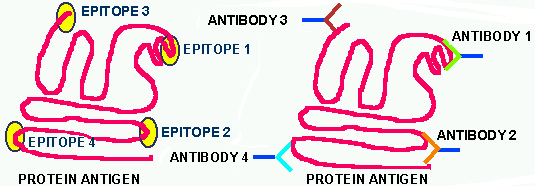
Figure 1. On the left is illustrated
a folded, functional protein. It might be an enzyme, or a cell wall receptor
site protein or a ribosomal protein etc. On this protein there are certain
groups
of amino acids that comprise epitopes. These groups are
defined as epitopes because they elicit an immune response
and for
no other reason. Since each of the epitopes is a different and unique
chemical cluster, each one of them induces a unique antibody. Each
antibody will bind tightly to its particular epitope and not to any
of the others. Within this cartoon lies the core information one needs
to understand how the immune system works. That is, if you know how one
car works, you have the core information on how all cars work, only some
details differ.
INNATE IMMUNITY = This can best be described
as GENETIC IMMUNITY or that immunity an organism is BORN WITH.
This type of immunity can be an immunity that applies to the vast majority
of the members of a species (SPECIES IMMUNITY), or it can be an
immunity that applies to only a certain subgroup within a species down
to a few individuals within that species. For example, cattle suffer from
the cowpox virus, but appear to have a SPECIES IMMUNITY to the closely
related smallpox viruses, whereas smallpox is a deadly disease to humans
, but cowpox is a mild localized skin infection. Humans are susceptible
to the HIV virus, but most of our related primates are immune to HIV, but
they suffer from HIV-like viruses to which we appear to be immune. Within
a species there may exist SUBGROUPS that are STATISTICALLY immune
or resistant to particular pathogens. For example, the Northern Europeans
appears to be more resistant to tuberculosis than are most Africans, whereas
Africans are naturally resistant to a variety of African diseases that
readily kill the "whites". Finally, because of the genetic variation within
every species INDIVIDUALS are statistically more resistant to some diseases,
and more susceptible to other diseases. Most of you know those within your
own families that "rarely" get colds or the flu, while other family members
catch one respiratory infection after another. While there are many factors
(diet, stress etc.) that could explain these individual differences, one
of them is that certain COMBINATIONS OF GENES render some more resistant
to the common cold viruses, whereas others of us are very susceptible.
This type of immunity has NOTHING TO DO WITH the type of specific immunity
we are discussing in this section.
ACQUIRED IMMUNITY = This refers
to immunity that one acquires in one of two ways, active or passive.
These are subdivided into the following further categories:
a) ACTIVE NATURALLY ACQUIRED IMMUNITY =
This occurs when individuals suffer from
a natural infection of a pathogen
and become immune to that pathogen upon recovery (e.g.
chickenpox)
b) ACTIVE ARTIFICIALLY ACQUIRED IMMUNITY
= This occurs when individuals are
actively vaccinated with an
antigen that confers immunity.
c) PASSIVE NATURALLY ACQUIRED IMMUNITY
= This occurs when individuals receive
antibodies from their mother
by a natural process, such as in BREAST MILK or in-utero transfer of
antibodies from mother to
fetus. In mammals, mother's milk is know to contain a large concentration
of antibodies and other antiviral
and antibacterial substance that protect the newborn infants. Further,
the mother's antibodies cross
the placental barrier, particularly near the end of term. In both these
circumstances the infant is
only resistant to whatever the mother is resistant to.
d) PASSIVE ARTIFICIALLY ACQUIRED IMMUNITY
= This occurs when individuals are
injected with POOLED serum
from immune individuals that contain antibodies against a large number
of pathogens. In the case
of humans, a fraction of blood serum, GAMMA GLOBULIN, that is
highly enriched in antibodies
is injected into individuals that have been exposed to certain pathogens.
The GAMMA GLOBULIN is obtained
from pooled sera from many individuals and thus contains a
broad spectrum of antibodies.
-
Gamma globulin is the material was given to people
who have been exposed to the hepatitis A virus in the contaminated strawberries
in the Spring of 1997.
LENGTH OF IMMUNITY
PASSIVE acquired immunity is short lived as
the antibodies eventually die off or are themselves removed from the body
as foreign protein. Since the person receiving the passive dose DOES NOT
PRODUCE their own antibodies, the immunity is TRANSIENT.
The ACTIVE forms of immunity are generally
long lived, particularly in the case of recovery from a CLINICAL INFECTION.
Sometimes this immunity it lifelong, but in other cases it is not. Vaccinations
may induce long-lived immunity, but recent data indicate that vaccinations
may not last as long as once was hoped. For example, there is a very effective
vaccine against tetanus, but it lasts only a few years and every year hundreds
of people who have been vaccinated against this bacterium die because they
have not gotten their BOOSTER SHOTS (vaccinations given periodically
to booster the immunity of previous vaccinations) every three to five years.
LEVELS OF IMMUNE SYSTEM
The specific immune system exists throughout the
body, but a major portion of it circulates in the blood and lymphatic systems,
as they flow throughout the body. The human specific immune system is a
two level or DUAL SYSTEM consisting of soluble antibodies and special
immune cells. The two systems work intimately as a coordinated unit. Foreign
material is dealt with by both components of this dual system. The cellular
components of the specific immune system includes a host of specialized
cells; new ones are being discovered all the time. The entire process of
specific immunity is initiated by non-specific immune cells, the phagocytic
cells of the nonspecific defense system, which act as general scavengers
and a kind of "attack dogs". These cells engulf or ingest any material
they perceive as foreign/nonself. Once inside these phagocytic cells the
engulfed material is digested and its chemical components are processed
for use by the specific immune system. The two components of the specific
immune system are described in greater detail below.
THE TWO COMPONENTS OF THE SPECIFIC
IMMUNE SYSTEM
I. THE HUMORAL IMMUNE SYSTEM.
One part of the duel level specific immune system
is called the HUMORAL system. The humoral system involves the soluble
ANTIBODIES described above. These antibodies circulate through the blood
and lymph system. When blood is spun in centrifuge, the red blood cells
(RBC) fall or PELLET to the bottom of the tube, leaving behind a
straw-colored
liquid called the SERUM. The antibodies are located in the
blood serum. Antibodies are made by SPECIAL B-CELLS, called PLASMA
CELLS that make and excrete antibody molecules.
II. THE CELL-MEDIATED IMMUNE
SYSTEM.
The second component of the specific immune system
involves a special class of cells called T-cells. There are several
important types of T-cells, each with unique responsibilities in immunity.
The T-cells do not produce antibody, but they react directly with other
cells. They might be thought of as the HIT-MEN of the immune system; point
out a foreign cell and they gang up on it, beat the dickens out of it until
there is nothing left but a few bit 'n pieces of garbage floating around.
Once in while they go crazy and decide to attack their own host cells and
then there is a serious problem (e.g. arthritis).
III. THE PHAGOCYTIC COMPONENTS OF
THE IMMUNE SYSTEM.
Cells of the nonspecific defense system, known
as
macrophages, monocyte and neutrophil, are involved in
a complex relationship in which they recognized and then ATTACK FOREIGN
MATERIAL, destroy it and process it for use by the specific immune
system. They use chemical signals to each other to coordinate their defense
of the host.
IV. COMPLEMENT.
Another important component of the specific immune
system is a group of proteins called the COMPLEMENT SYSTEM. Complement
is a GROUP OF PROTEINS that, like the antibodies, are soluble and
reside in the serum. Complement is a COMPLEX OF ENZYMES that mainly act
on foreign cells by punching holes their membranes to cause their LYSIS
AND DEATH. Complement works in concert with the SPECIFIC ANTIBODIES
that "point out" the cells to be attacked by the complement; i.e., the
antibodies act to "FINGER" (identify) a target cell and the complement
acts as the "HIT MAN" that kills the targeted cell. In addition
complement, plus antibody, designate which cells are to be engulfed by
the phagocytic cells. Complement can also result in immunological damage
to ones own cells in the case of diseases caused by faulty immune systems.
One such reaction is the serious allergic response known as ANAPHYLACTIC
SHOCK.
DEVELOPMENT OF THE IMMUNE SYSTEM
Now we come to issue brought up in the introduction,
namely: How does the body distinguish the good-guys from the bad-guys?
The steps in the immune system development are:
a) Stem cells, which are the PARENT CELLS
of all immune cells, enter the liver of the fetus and
develop to a point there.
b) From the liver some stem cells move into the
bone marrow (at the center of the bones) where
they differentiate into B
CELLS and NATURAL KILLER CELLS.
c) Other stem cells move from the liver into
the thymus gland located in the middle of your chest.
d) The thymic stem cells differentiate in a variety
of T cells.
e) Other stem cells go on to differentiate into
other blood cell lines such as macrophages.
The immune system is spread throughout the entire
body and includes the following (a partial listing):
-
Adenoids & tonsils
-
The lymph nodes & lymphatic system
-
The Spleen, appendix and small intestine
-
The bone marrow
-
The thymus gland
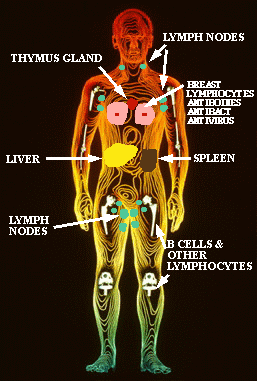 Figure
4. This figure shows the location in the body of various components of
the nonspecific and specific immune systems. The B cells and a variety
of other lymphatic cells are made in the bone marrow. The lymph nodes contain
the macrophages, B cells and T cells, which is why your lymph glands swell
up and become tender to the touch when you have an infection. The thymus
gland is the gland where the differentiation of the T cells occurs. Other
macrophages, monocytes and phagocytes reside in the liver, spleen and lungs.
Special immune cells have been found in the brain, in the skin and in the
cells lining the intestine. Breast milk contains a variety of the mother's
white blood cells that kill microbes in the infant's gut and stimulate
the development of the infant's immune system as well as antibodies and
10 other microbial inhibitors (Sci. Am. Dec. 1995)
Figure
4. This figure shows the location in the body of various components of
the nonspecific and specific immune systems. The B cells and a variety
of other lymphatic cells are made in the bone marrow. The lymph nodes contain
the macrophages, B cells and T cells, which is why your lymph glands swell
up and become tender to the touch when you have an infection. The thymus
gland is the gland where the differentiation of the T cells occurs. Other
macrophages, monocytes and phagocytes reside in the liver, spleen and lungs.
Special immune cells have been found in the brain, in the skin and in the
cells lining the intestine. Breast milk contains a variety of the mother's
white blood cells that kill microbes in the infant's gut and stimulate
the development of the infant's immune system as well as antibodies and
10 other microbial inhibitors (Sci. Am. Dec. 1995)
CLONAL SELECTION
Consider the problem an immune
system faces. It must defend its host against thousands of unknown , each a MOSAIC of different antigens (epitopes).
Further, it must distinguish between millions of self antigens and other
millions of foreign antigens; the penalty for failure is DEATH by
a pitiless nature. What makes this goal even more difficult is that many
of the self-antigens are chemically very similar to the nonself-antigens.
As the early immunologists defined this incredible diversity they were
awestruck and puzzled as to how this could possible be. It was one of these
situations that was demonstrability true, but seemed impossible to achieve;
but then life itself fits in that category doesn't it? As usual in science
the answer came from the brilliant reasoning of a few people. The thought
process that broke the "case" went something like this.
Instead of thinking that the immune system had
to be INSTRUCTED AHEAD OF TIME as to which antibodies would be required
throughout a life time, clearly an impossible task, N.K. JERNE suggested
that the immune system was SELECTIVE rather than instructive. Jerne
reasoned that the immune system must randomly made billions of different
specific-epitope-binding
antibodies and then let the antigens that accidentally stumbled into
the host chose or select which antibodies would be produced in quantities
large enough to be protective. In a sense this is just another twist on
the "survival of the fittest" process in evolution. Burnet in Australia
and Talmage in CO then hypothesized that antibodies
SIT ON THESURFACE
of lymphocytes and that each lymphocyte manufactures only a SINGLE ANTIBODY
(which recognizes and binds to only a SINGLE epitope). This theory
has been shown to be essentially correct by a number of brilliant experimentalists.
THE CURRENT THEORY OF ANTIBODY FORMATION:
1. During fetal development the body randomly produces
millions of B & T CELLS, each of which produces only a .
2. The B cells that produce self antibodies are
DESTROYED,
leaving only lines or CLONES of B cells that produce random antibodies
to foreign epitopes.
3.When a particular foreign epitope (for
example, antigen 2,025) appears in the host's body it is PROCESSED
by lymphocytic cells of the nonspecific defense system. This sets off a
sequential series (cascade) of events that eventually acts on a small population
of randomly-produced B/T cells that happen (by chance) to have on their
surface, antibody (2,025) which binds to ANTIGEN 2,025.
4. These events trigger a RAPID PROLIFERATION
of that PARTICULAR B (and T-cell) cell population (2,025),
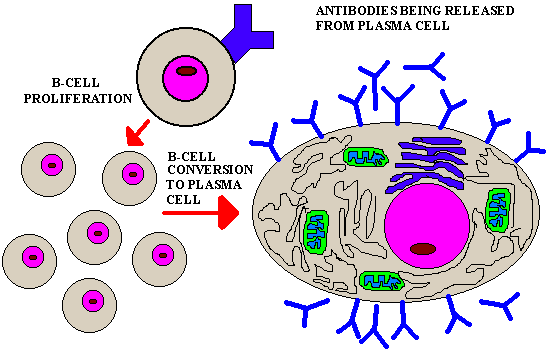
producing a large number of clones. These 2,025 B cell-clones differentiate
into PLASMA CELLS (Fig. 3) which are ANTIBODY-PRODUCING-FACTORIES
that spew out prodigious quantities of ONE ANTIBODY-#2,025, that
binds to the specific antigen-epitope 2,025 that stimulated it.
5. The specific antibody floods through the host
and wherever it binds to its epitope it MARKS IT FOR ATTACK and
destruction by the appropriate cells and associated components of the immune
tem (complement and PMNs etc.).
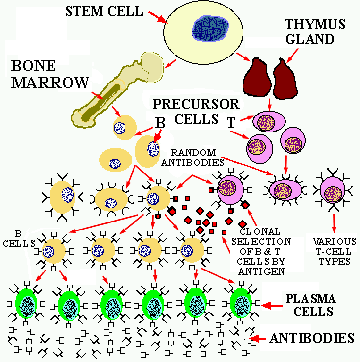 Figure
3. The process of B & T cell differentiation and CLONAL SELECTION.
The parental STEM cells migrate to the bone marrow and to the thymus gland
where they differentiate into B and T cells which make random epitope binding
proteins. When a foreign epitope binds to the appropriate site on the B
& T cells, they replicate into clones that, in the case of the B cells
differentiate into PLASMA cells that produce prodigious quantities of specific
antibodies. The T cell clones further differentiate into several different
T cell types with specific functions.
Figure
3. The process of B & T cell differentiation and CLONAL SELECTION.
The parental STEM cells migrate to the bone marrow and to the thymus gland
where they differentiate into B and T cells which make random epitope binding
proteins. When a foreign epitope binds to the appropriate site on the B
& T cells, they replicate into clones that, in the case of the B cells
differentiate into PLASMA cells that produce prodigious quantities of specific
antibodies. The T cell clones further differentiate into several different
T cell types with specific functions.
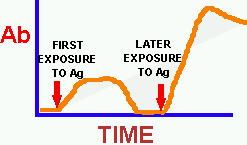 Figure
4. The response to an antigen (Ag) in terms of the production of a specific
antibody over time. Initially the levels of each unique antibody are extremely
low, however as soon as the stimulation events occur (Fig. 7) and the plasma
cell clone begins producing antibodies the TITER (concentration
or quantity/volume) of the unique antibody begins to rise. It takes about
2 weeks for the Ab level to peak. Once the foreign antigen is removed,
antibody production slowly returns to a low level, however MEMORY PLASMA
CELLS remain in the system. When the original antigen again appears
in the host these memory cells respond rapidly and produce even higher
levels of antibodies. This "REMEMBERING RESPONSE" is why we remain
immune to many diseases for a long time. The secondary exposure to the
antigen may be natural or it may be artificial in the case of booster vaccinations.
As parents we are responsible for seeing to it that our children are initially
vaccinated and that their booster shots are given at the appropriate ages.
Figure
4. The response to an antigen (Ag) in terms of the production of a specific
antibody over time. Initially the levels of each unique antibody are extremely
low, however as soon as the stimulation events occur (Fig. 7) and the plasma
cell clone begins producing antibodies the TITER (concentration
or quantity/volume) of the unique antibody begins to rise. It takes about
2 weeks for the Ab level to peak. Once the foreign antigen is removed,
antibody production slowly returns to a low level, however MEMORY PLASMA
CELLS remain in the system. When the original antigen again appears
in the host these memory cells respond rapidly and produce even higher
levels of antibodies. This "REMEMBERING RESPONSE" is why we remain
immune to many diseases for a long time. The secondary exposure to the
antigen may be natural or it may be artificial in the case of booster vaccinations.
As parents we are responsible for seeing to it that our children are initially
vaccinated and that their booster shots are given at the appropriate ages.
ANTIBODY STRUCTURE AND VARIABILITY
So at this point we know that there are millions
of B-cell-antibody-producing types, just waiting to be "triggered" by contact
with their respective antigen, but we still don't know how we get
these millions of different B-cells in the first place. To understand
how this occurs you have to know something about antibody structure.
There are five different types of antibodies,
however in this course we will discuss only the most common one, IgG,
in detail. However, note that the other 4 types physically resemble the
basic structure of IgG. IgG does most of the humoral immune work. The figure
below shows the physical structure of the IgG molecule.
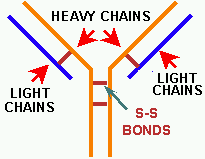
Figure 5. The IgG molecule. IgG
is composed of two protein subunits, a LIGHT (blue) and a HEAVY
CHAIN (orange) named appropriately according to their relative sizes.
The various chains are bonded together to form the IgG molecule with disulfide
bonds (S-S bonds). Note the two arms & the heavy 'n light chains.
The Y-shaped structure is real as electron microscopic
pictures show. However, even before they viewed IgG in an electron microscope
immunologist had discerned its basic shape. They knew that each antibody
had to have two equivalent binding sites for its specific epitope. It turns
out that those two binding sites are located at the end of the short arms
of the Y.
The IgG molecule is further divided into
CONSTANT
and VARIABLE REGIONS OR DOMAINS. The constant regions have mostly
the SAME amino acid sequence in all IgG molecules (we won't discuss the
differences here), whereas the amino acid sequences in the variable regions
are DIFFERENCE for
produced by a clone of plasma cells. The amino acid sequence in the variable
domains are such that they tightly bind to particular epitopes. Thus they
show the same lock-key relationship as do enzymes/substrates
and enzymes/allosteric molecules and viruses/target cell receptors.
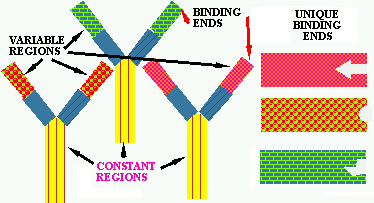 Figure
6. Three unique antibody IgG molecules. The base of the "Y" and part of
each arms are called the CONSTANT REGIONS because their amino acid sequence
tends to be very similar in all IgG molecules. The variable regions are
at the end of the arms and their amino acid sequence is very different
for each IgG molecule. These variable regions fold so as to bind to specific
epitopes or antigens; the unique binding sites are shown in their respective
three variable regions on the right.
Figure
6. Three unique antibody IgG molecules. The base of the "Y" and part of
each arms are called the CONSTANT REGIONS because their amino acid sequence
tends to be very similar in all IgG molecules. The variable regions are
at the end of the arms and their amino acid sequence is very different
for each IgG molecule. These variable regions fold so as to bind to specific
epitopes or antigens; the unique binding sites are shown in their respective
three variable regions on the right.
ANTIBODY VARIABILITY
Antibody variability comes about through an unusual
SHUFFLING
of the genes that code for the variable portions of the IgG molecule. The
antibody genes are inherited as GENE FRAGMENTS. During lymphocyte
development these gene fragments are joined together in RANDOM ARRANGEMENTS
it form the COMPLETE GENES in the individual B & T cells. The
fact that the IgG molecules are composed of two proteins, each with its
independently produced variable regions adds increased variability to the
whole process. It is estimated that >100 million distinct antibodies can
be made by this process. In addition the genes for receptors of B lymphocytes
MUTATE extremely rapidly when the B cell is activated by binding to a foreign
substance or antigen. Once a B lymphocyte binds antigen to its receptor,
it differentiates and secretes specific antibody molecules that have been
specified by the genes that created the receptor on the parent B cell.
TYPES OF ANTIBODY (Ab) REACTIONS
The basic reaction of all Ab's with their epitopes
is the same (a binding of ligand and receptor), but the physical
MANIFESTATIONS
of that reaction differs depending on the PHYSICAL NATURE of the
antigen. The point to remember is that the Ab has TWO binding sites so
a single Ab molecule can bind to two independent antigen molecules or particles.
You see one these manifestations in the lab.
NEUTRALIZATION = When the antigen is a soluble
toxin, the addition of an Ab against it will usually render the toxin INEFFECTIVE
(nontoxic), that is it NEUTRALIZES it. Such neutralized toxins are
called TOXOIDS and can be used as vaccines. For example, if you
were suspected of suffering from either tetanus or botulism poisoning the
treatment would involve giving you a shot of the appropriate antitoxin,
which is a common name for the Ab against a toxin. The antitoxin circulates
through your body and binds and neutralizes any toxin it contacts.
PRECIPITATION = Under the proper conditions
a soluble antigen can be precipitated in the presence of its Ab because
of the of antigen-antibody net-work that forms gets large enough to form
masses that SETTLE OUT (precipitate) on their own.
AGGLUTINATION = When the antigen is a large
PARTICLE,
like a whole bacterium or a RBC, the addition of its Ab will form an Ab-antigen
net work that causes the particles to CLUMP IN LARGE MASSES like milk coagulating
when it spoils. This agglutination is easy to see and is useful for diagnostic
purposes. For example, if you want to see if a person is making Ab against
a particular bacterium, mix the person's serum with the suspected bacterium;
if the bacteria clump into large globs it means that Ab are present. Both
precipitation and agglutination are illustrated below.
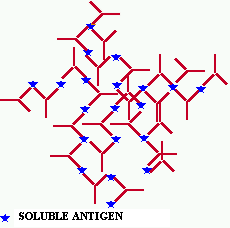 Figure
7. Antibody/antigen complex forming a larger complex. These nets can grow
so large that they become insoluble and visible to the eye. The network
forms because of the dual-binding characteristic of the antibody which
allows it to attach to two different antigen molecules at the same time.
Figure
7. Antibody/antigen complex forming a larger complex. These nets can grow
so large that they become insoluble and visible to the eye. The network
forms because of the dual-binding characteristic of the antibody which
allows it to attach to two different antigen molecules at the same time.
T-CELL IMMUNITY
The second component of the adaptive immunity system
involves a set of special immune cells called T cells. We will only
deal with three of the T cell types. The T cells develop in the thymus
gland, but the process is not completely understood. Briefly, the stem
cells in the thymus undergo differentiation's that form two major groups
of T cells, the KILLER T CELLS (Tc or Tk) and the HELPER T CELLS
(Th). The process of immunological diversification through DNA fragment
shuffling is the same as that which was described for the B cell development
so that EACH Tk and Th cell responds only to a unique epitope. T cells
that react with self antigens DIE OFF during the early stages of differentiation.
The T cell clones migrate throughout the lymphatic system. When a T cell
encounters its antigen (epitope) it goes through a series of changes that
convert it into its final immunological defense posture.
THE SPECIAL ROLE OF THE T-HELPER (Th)
CELL
The special role of
the T-helper (Th-cell) in developing immunology is described below:
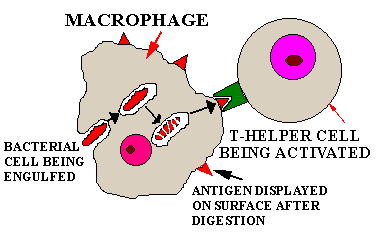
-
A macrophage engulfs a virus or bacteria & breaks
down their proteins. Antigenic fragments of these proteins are presented
on the surface of the macrophage.
-
The few Th-cells, which contain receptors
on their surface that recognizes a particular presented foreign antigen,
INTERACTS
WITH that unique-foreign antigen. This interaction triggers a series
of events that ACTIVATES the Th-cell.
-
The activated Th-cells are stimulated to proliferate,
producing a population of this class of Th-cells (clonal selection). The
activated Th-cells interact physically (have contact with) with only
those rare B-cells that make antibody that recognizes the SAME antigen
molecules that have activated the Th-cell. During this interaction the
Th- & B-cells recognize each other
-
by their common recognition of the unique antigen.
-
The B- and Th-cell interaction stimulates the Th-cells
to produce chemicals (CYTOKINES) that, in turn, stimulate the appropriate
B-cells to proliferate (clonal selection) and to DIFFERENTIATE into
Ab-producing plasma cells that produce the Ab that bind the antigen that
the Th-cell originally reacted to.
Therefore, the Th helper cell acts as a MASTER
CONTROL CELL of the immune system. It is REQUIRED for
both the humoral and cellular immune systems to function. When Th cells
are
not present the host's fate is sealed because the correct B-cells
will not proliferate and the correct antibody will not be produced (like
a football team without a quarterback) with the result that death ensues.
THE FUNCTION OF T-KILLER (Tc) CELLS.
The T killer cells (Tc) have a different function.
The Tc cells are designed to recognize foreign antigens on the SURFACE
OF HOST CELLS. Foreign cell epitopes appear on host cells mainly in
two types of situations, in viral infection and in cancer cells. In both
these case there are changes in the composition of the host's cells that
cause foreign antigens to be PRESENTED ON THE SURFACE of the modified
cell. The Tc cells recognize these foreign epitopes and are stimulated
to attack and destroy the infected or modified (e.g. cancer) cell.
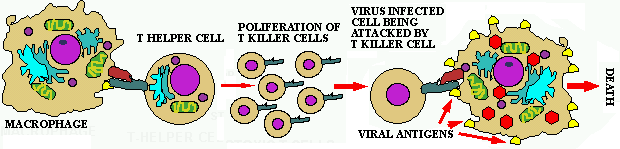
Figure 8. Activation &
killing by Tc killer cells of cells displaying a unique surface antigen.
Note the virus particles in the cell on the right and the presence of unique
viral proteins on its surface to which the Tc cells bind.
Other T cell types exist and probably more types
will be found. The above is an incomplete and simplified explanation of
what is currently known about the immune system. Some of it will undoubtedly
be modified as new facts come to light and we will surely find that it
is even more complex and subtle than previously imagined. It's like human
relationships which usually start out simple, but the become more complex
as time goes on.